第一作者:Tom Wieczorek
通讯作者:Jinqiao Dong(董金桥),Anthony P. Davis
通讯单位:上海交通大学,布里斯托大学
DOI: https://doi.org/10.1016/j.chempr.2026.103068
- 研究背景
人们长期关注如何利用分子,尤其是序列明确的聚合物,来实现数字信息的存储、读取和处理。原则上,任何由特定单体序列构成的聚合物都可以被视为一种信息载体;但在实际体系中,只有当这些信息能够在分子水平上被选择性读取、复制并传递时,才真正具备信息功能。核酸为这一目标提供了最典型的范例:DNA 和 RNA 不仅能够储存遗传信息,还能通过碱基互补配对实现信息复制、翻译和程序化自组织,从而支撑遗传、生命繁殖和达尔文进化等基本生命过程。然而,核酸未必是唯一能够承担信息存储与传递功能的分子体系。寻找核酸之外的信息聚合物,不仅有助于发展新型可编程分子材料,也可能为理解生命起源提供新的线索。在前生命地球环境中,从随机分子混合物演化为具有特定功能的生命体系,可能需要早期形式的达尔文进化参与。尽管 RNA 世界假说认为 RNA 可能早于 DNA 成为遗传物质,但如此复杂的分子在生命最初阶段即大量出现并不一定容易。因此,一个重要科学问题是:在 RNA 接管遗传功能之前,是否存在更简单、更易形成的聚合物体系曾经承担过早期信息存储和传递的功能?
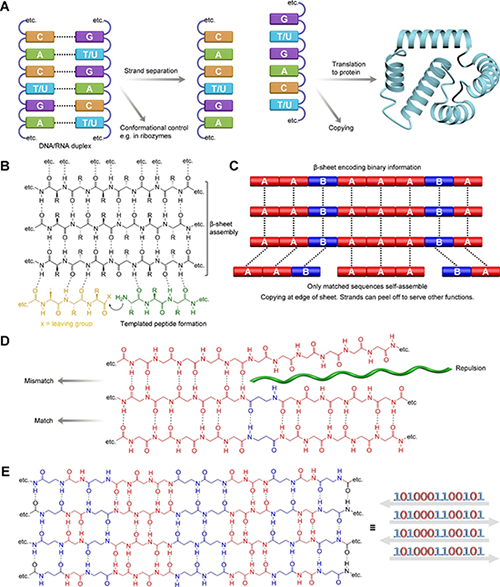
图1. 分子摩尔斯密码(MMC)是利用α-氨基酸与β-氨基酸主链长度差异作为二进制编码单元,通过 β-折叠中的氢键匹配实现肽序列信息读取、选择性自组装与潜在信息传递的分子编码策略。
- 研究问题
多肽和类肽分子为这一问题提供了重要研究对象。酰胺键易于形成且稳定,使肽类分子在前生命化学中具有较高可信度。尤其是肽的 β-折叠结构,由于能够通过氢键在片层边缘模板化组装新的肽链,被认为具有潜在的自我复制能力。与传统双链复制体系不同,β-折叠中新生成的链本身可以继续作为模板,因此理论上无需链分离即可实现连续复制。然而,β-折叠体系虽然具有模板复制潜力,却并不天然具备明确的信息编码能力。要实现类似核酸的信息功能,肽链必须能够容纳不同结构单元,并且这些单元的排列顺序能够被体系中其他分子选择性识别和传递。传统肽可以通过侧链序列编码信息,但这种信息如何在 β-折叠组装中被可靠读取和复制,仍然缺乏清晰机制。针对这一挑战,研究者提出了“分子摩尔斯密码”(molecular Morse code, MMC)的概念。其核心思想是利用 α-氨基酸和 β-氨基酸在主链长度上的差异,将“短”和“长”的结构单元作为分子层面的编码比特(如图-1所示)。β-氨基酸插入 β-折叠结构后,会改变主链 NH 和 CO 基团的空间排列,使不匹配的肽链发生氢键错位;而当另一条肽链在相应位置也含有匹配的 β-氨基酸时,则能够恢复规则的反平行 β-折叠配对。由此,α/β 氨基酸序列可以像摩尔斯密码中的点和划一样编码信息,并通过氢键匹配实现分子水平的选择性读取。
- MMC对多肽折叠构象的调控
本研究的目标是证明,基于MMC的分子内缔合作用可以用于调控α/β-肽的折叠构象。研究人员制备了20条肽分子,这些肽在DPro-Gly转角诱导单元两侧含有等长的 α/β序列。其结构如图2所示,这些肽保留了固相合成过程中引入的 N-Fmoc 端基,并在C端通过甲醇酯化。它们被设计用于有机介质中的研究,以确保MMC调控所需的氢键作用能够有效发挥。对于识别序列主要采用具有较大侧链、且已知有利于 β-折叠形成的氨基酸。其中,L-缬氨酸和 L-亮氨酸被用作 α-氨基酸,(S)-β³-高苯丙氨酸(β³hPhe)和 (S)-β³-高缬氨酸(β³hVal)被用作β-氨基酸。在少数情况下,也使用β-Ala作为 β-氨基酸。预期是:当这些序列在 MMC 意义上匹配时,能够形成 β-折叠;而当序列错配时,则会以其他构象为主。
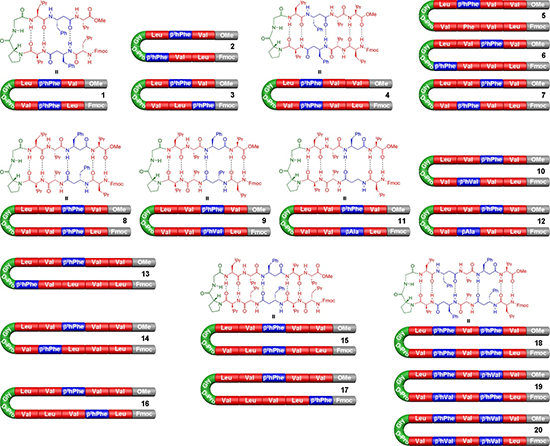
图2. MMC介导的多肽折叠调控。用于研究MMC构象调控的肽序列1–20:α-氨基酸以红色表示,β-氨基酸以蓝色表示,促进β-转角形成的DPro-Gly 单元以绿色标出
多肽1–20通过标准固相合成方法制备。然而,部分肽的溶解性成为研究中的一个挑战。十肽6极难溶解,因而无法开展研究;若干十二肽也难以纯化,只能获得少量样品。进一步研究十四肽的尝试也因溶解性问题受阻,因此最终研究对象限制在12个残基以内。研究者通过圆二色谱(CD)在甲醇中考察这些肽的二级结构,并在许多情况下利用二维核磁共振(2D NMR)进行验证。在肽的CD光谱中,反平行β-折叠结构通常表现为 218 nm 附近的负峰和 195 nm 附近的正峰。CD光谱为MMC假说提供了有力支持。如图3所示,匹配与错配结构之间表现出明显差异。图3A中,匹配的十肽4和8在198 nm处呈现强正峰,并在223 nm附近出现负峰,接近β-折叠的典型光谱特征。相比之下,错配结构5和7的光谱特征较弱,说明其主要为无序结构。进一步地,肽4和8在CDCl₃中的2D NOESY NMR 光谱显示出多个链间空间相关信号,支持其形成 β-发夹构象。相反,肽5和7的ROESY光谱仅显示链内相关信号,进一步说明其未形成有效的 β-发夹结构。多组对比进一步证明,在一定范围内改变侧链并不会破坏 MMC对折叠的调控。例如,匹配序列9和11与8类似,给出了与β-发夹结构一致的数据;而错配序列10和12则表现为无序结构。值得注意的是,肽11和12中含有最简单的β-氨基酸β-Ala,说明MMC策略并不依赖于复杂β-氨基酸侧链。
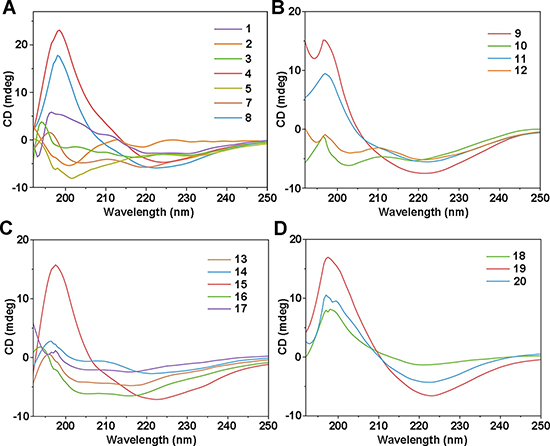
图3. 圆二色谱证实 MMC 介导多肽折叠的调控
图3C展示了十二肽13–17的CD光谱,这是一组β³hPhe 扫描实验,即逐步将 β³hPhe移向N端。结果同样显示出清晰区别:只有匹配序列15表现出符合β-发夹结构的 β-折叠光谱特征,其余序列则主要为无序构象。匹配的十二肽18–20也均表现出预期的β-折叠CD特征,说明体系中可以容纳不止一个β-氨基酸,同时进一步证明了侧链类型具有一定可变性。因此结果清楚表明:MMC 确实可以用于以理性、可编程的方式调控肽分子的构象。
- MMC调控多肽可编程组装
为进一步验证MMC的作用,研究者将其拓展到水相 β-折叠聚集体的自组装体系。受 MAX系列发夹肽启发,他们设计了全α-肽21以及 α/β-肽 22、23 和 24(图4),通过 pH 调控赖氨酸去质子化,诱导肽分子由可溶状态转变为 β-折叠聚集体并形成自支撑水凝胶。CD 和 ATR-FTIR 光谱表明,肽21–24在酸性条件下主要为无序构象,而在pH 11下转变为β-折叠结构。TEM进一步显示,不同MMC编码的肽形成了可区分的纤维形貌:全α-肽21形成缠结单纤维网络,平均直径约5.24 nm;α/β-肽22形成较有序排列的纤维,平均直径约4.52 nm;α/β-肽23形成特征性扭曲带状结构;α/β-肽24则形成非扭曲多股带状结构。为验证自分类,研究者采用两种混合方式:先混合后自组装(M-SA)和先自组装后混合(SA-M)。在21+23和21+24体系中,两种方法得到的TEM图像高度相似,并均可清楚观察到两类独立聚集体(图5和图6),说明不同 MMC 编码的肽并未随机共组装,而是倾向于与相同编码的肽自组装。这些结果表明,MMC不仅能够控制分子内多肽构象折叠,还能够在水相中调控分子间 β-折叠自组装与自分类。其核心意义在于,α/β氨基酸序列可作为可读取的分子编码,引导不同肽分子选择性识别“同类”,并程序化形成结构可区分的水凝胶纤维。
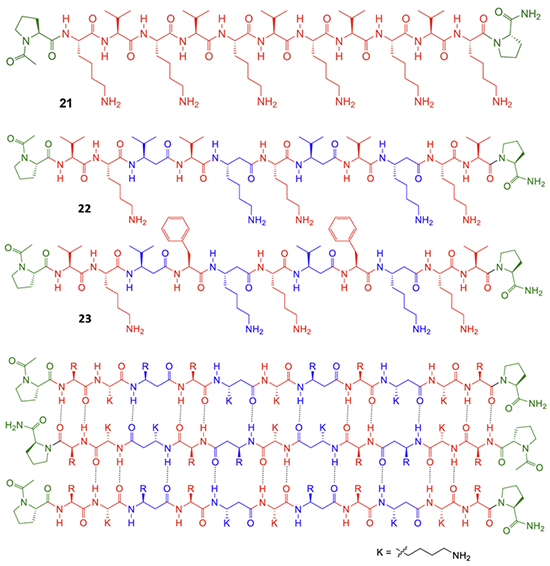
图4. 多肽21-23的设计与合成
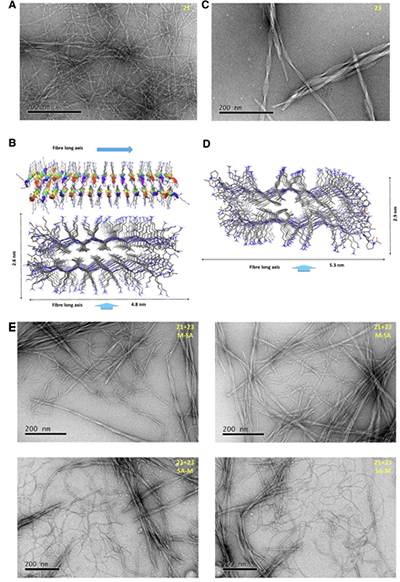
图5. MMC编码可驱动21和23在水相中选择性自分选,形成两类独立纤维
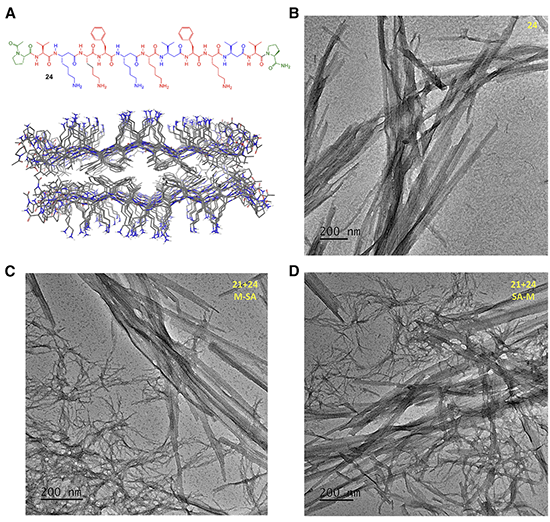
图6. MMC编码可驱动21和24在水相中发生自分类组装
总结与展望
DNA 的关键信息功能包括三个方面:可延伸的重复聚合物结构、可变组分形成的信息编码能力,以及通过互补识别实现模板导向复制。在结构上远离核酸的合成聚合物中复制这些特征一直很困难。本文提出的分子摩尔斯密码(MMC)表明,反平行β-折叠体系至少具备部分类似DNA的信息存储与传递能力。通过α-氨基酸和β-氨基酸的“短/长”主链差异,肽链序列能够以可预测方式进行选择性配对,从而控制分子构象、自组装和自分选。该体系由化学简单、可变性强的氨基酸组分构成,原则上可承载多样化序列信息。与DNA不同,MMC体系形成的是延展聚集体,而非离散双链结构。因此,它的潜在应用更可能与核酸体系互补,而不是替代核酸。例如,本文展示的混合自分选水凝胶即体现了MMC在程序化超分子材料中的应用潜力。从生命起源角度看,MMC也具有启发意义。虽然目前不能断言 MMC 曾参与生命起源,但在“淀粉样世界”假说中,β-折叠肽可能具有复制和催化功能,而其信息存储机制尚不清楚。MMC为这一问题提供了一种可能答案:α-和 β-氨基酸结构简单,具有前生命化学可信度,并且β-折叠聚集体有可能无需链分离即可实现连续复制。总体而言,本文证明了MMC可作为一种储存和传递序列信息的分子编码策略,并可用于调控肽的折叠构象和自组装。尽管基于MMC的信息自我复制尚未实现,但已有α-肽自复制研究表明这一目标具有可行性。如果未来实现,MMC体系可能为构建能够自主、连续复制并发生达尔文进化的人工分子系统提供新路径。